



Bacopa (Bacopa monnieri L.)
 Gli effetti benefici della Bacopa (Bacopa monnieri L.) sulla memoria e sulla cognizione sono stati ampiamente studiati e molti articoli di revisione scientifica hanno descritto le sue proprietà nootropiche [1-4]. L’attività della Bacopa è stata attribuita al suo contenuto di saponine, principalmente bacosidi A e B [5]. Di queste specie chimiche, quella nota per essere responsabile delle sue proprietà nootropiche è il bacoside A. Secondo vari studi, è stato suggerito che la Bacopa possiede proprietà neuroprotettive e migliora le capacità cognitive, grazie alla sua capacità di modulare il sistema colinergico e di combattere il processi responsabili del danno ossidativo nel cervello [6-9]. Il cervello è particolarmente suscettibile al danno ossidativo, a causa della sua elevata attività metabolica, dei suoi alti livelli di ferro, che è una specie pro-ossidante (che induce danno ossidativo) ed è composto da lipidi insaturi, inclini alla perossidazione [10].
Gli effetti benefici della Bacopa (Bacopa monnieri L.) sulla memoria e sulla cognizione sono stati ampiamente studiati e molti articoli di revisione scientifica hanno descritto le sue proprietà nootropiche [1-4]. L’attività della Bacopa è stata attribuita al suo contenuto di saponine, principalmente bacosidi A e B [5]. Di queste specie chimiche, quella nota per essere responsabile delle sue proprietà nootropiche è il bacoside A. Secondo vari studi, è stato suggerito che la Bacopa possiede proprietà neuroprotettive e migliora le capacità cognitive, grazie alla sua capacità di modulare il sistema colinergico e di combattere il processi responsabili del danno ossidativo nel cervello [6-9]. Il cervello è particolarmente suscettibile al danno ossidativo, a causa della sua elevata attività metabolica, dei suoi alti livelli di ferro, che è una specie pro-ossidante (che induce danno ossidativo) ed è composto da lipidi insaturi, inclini alla perossidazione [10].
Il sistema colinergico è costituito da cellule nervose organizzate che utilizzano il neurotrasmettitore acetilcolina nella trasduzione del segnale. Queste cellule producono, rilasciano o vengono attivate dall’acetilcolina durante la propagazione dell’impulso nervoso. Il sistema colinergico è stato associato a diverse funzioni cognitive come la memoria, l’attenzione selettiva e l’elaborazione delle emozioni [11].
Un’altra proprietà importante della Bacopa è la sua attività antinfiammatoria nel cervello [12]. La neuroinfiammazione, a lungo termine, è dannosa e può causare neurodegenerazione e può portare all’insorgenza di malattie come l’Alzheimer, il Parkinson e la sclerosi multipla [12]. Studi in vitro con cellule microgliali (cellule del tessuto nervoso) hanno dimostrato che gli estratti alcaloidi di Bacopa, così come il bacoside A, inibiscono il rilascio di citochine pro-infiammatorie come il fattore di necrosi tumorale alfa (TNF-α) e l’interleuchina 6 (IL-6) [13]. Questi composti erano anche correlati all’inibizione di enzimi associati all’infiammazione nel cervello [12].
L’estratto di Bacopa di MINFIRE è ottenuto da metodi di estrazione avanzati, in cui si ottiene un estratto con un alto grado di purezza e un’alta concentrazione di bacosidi. Per produrre solo 1 kg del nostro estratto 30: 1 e raggiungere la sua concentrazione minima del 50% in bacosidi, sono necessari 30 kg di Bacopa! La concentrazione del 50% di bacosidi nel nostro estratto è una delle più alte sul mercato, poiché normalmente gli estratti utilizzati in altri prodotti sono nell’ordine del 25% -45% in bacosidi. Inoltre, il nostro estratto soddisfa tutti i requisiti di qualità dell’industria degli integratori alimentari e i criteri molto esigenti dei prodotti Nutribiolite.
Hericium o Criniera di leone (Hericium erinaceus Bull. Pers.)

L’hericium, noto anche come criniera del leone, è un fungo distribuito tra il Nord America e l’Asia. Questo fungo ha una lunga storia nella medicina tradizionale cinese, a cui vengono attribuite proprietà benefiche per la salute del cervello [14]. Infatti, studi più recenti attribuiscono un grande potenziale all’hericio nel trattamento delle malattie neurologiche, poiché contiene composti neurotrofici bioattivi che possono passare attraverso la barriera emato-encefalica [15, 16]. Questo fungo è ricco di composti bioattivi come i polisaccaridi β-glucani e i terpenoidi hericenone ed erinazina.
I Β-glucani sono stati oggetto di particolare interesse per la loro bioattività. Ad esempio, studi in vivo hanno identificato gli effetti protettivi di questi sull’epatotossicità ossidativa nei topi [17]. Gli effetti neuroprotettivi del β-glucano sono stati osservati anche in studi in vitro con cellule tossiche a causa della formazione di placche β-amiloidi. Le placche Β-amiloidi sono depositi di proteine, che normalmente si verificano nel cervello e sono associati alla malattia di Alzheimer, soffocando le cellule cerebrali e contribuendo alla progressiva degenerazione dei neuroni. I polisaccaridi Β-glucano hanno anche promosso la vitalità cellulare e protetto le cellule contro l’apoptosi indotta dalla formazione di placche β-amiloidi [18].
L’erinacina della Hericium è una famiglia di terpenoidi, molecole organiche in grado di attraversare la barriera emato-encefalica. Il grande interesse per gli estratti di Hericium si basa principalmente sulla presenza di erinacina. È stato dimostrato in diversi studi scientifici che questa molecola è in grado di indurre la sintesi della proteina Nerve Growth Factor (NGF) [16, 19], essenziale per lo sviluppo e il mantenimento dei neuroni del sistema nervoso periferico e per l’integrità funzionale del neuroni colinergici (del sistema colinergico) presenti nel sistema nervoso centrale [20].
In MINFIRE il fungo Hericium (o criniera del leone) subisce un avanzato processo di estrazione con una miscela idroalcolica di etanolo: acqua per ottenere una maggiore quantità di molecole organiche solubili come l’erinacina. Per produrre solo 1 kg del nostro estratto sono necessari 10 kg di funghi di criniera del leone. Questo estratto è normalmente standardizzato in termini di polisaccaridi totali. Meno polisaccaridi saranno presenti, più composti organici come l’erinacina, motivo per cui il nostro estratto contiene il 30% di polisaccaridi, per avere il miglior rapporto tra β-glucano polisaccaridi ed erinacina. Altri estratti hanno più polisaccaridi, che diminuiranno il loro contenuto di erinacina.
Ginkgo (Ginkgo biloba L.)

Con l’età, il corpo soffre di una diminuzione dell’elasticità e del tono dei vasi sanguigni, che limita il flusso sanguigno. Il flusso sanguigno ridotto si traduce in una minore quantità di ossigeno e altri nutrienti nel cervello, il che può portare a una funzione cognitiva compromessa. L’estratto di Ginkgo biloba esercita effetti dilatanti e costrittivi sui vasi sanguigni, aumentando l’afflusso di sangue al cervello e, di conseguenza, i suoi livelli di glucosio e ATP, con conseguente miglioramento della funzione mentale [21]. La sperimentazione in vivo ha dimostrato che i suoi composti fitoattivi (glicosidi flavonoidi e lattoni terpenici) attraversano facilmente la barriera emato-encefalica, agendo direttamente nel cervello, producendo gli effetti benefici sopra menzionati [22]. Secondo questi studi, il suo meccanismo d’azione è attraverso la sua attività antiossidante e antipiastrinica (PAF), oltre ai suoi effetti modulanti sul tono cerebrovascolare, sull’attività recettore / trasmettitore, sul metabolismo del glucosio e sull’attività elettroencefalografica [22-26].
Come nel caso dell’estratto di Bacopa, studi recenti hanno suggerito che l’estratto di Ginkgo ha anche effetti diretti sul sistema colinergico aumentando la capacità dell’acetilcolina di legarsi a un sito recettore sinaptico, il che potrebbe spiegare i suoi effetti di potenziamento cognitivo sia acuto che cronico [27].
L’estratto di ginkgo standardizzato di MINFIRE contiene almeno il 24% di flavonoidi glicosidi e il 6% di terpeni lattoni. È uno degli estratti più apprezzati e utilizzati per la prevenzione della demenza e del deterioramento cognitivo. Per preparare 1 kg del nostro estratto vengono utilizzati 50 kg di foglie di Ginkgo secche.
Tè verde (Camellia sinensis L. Kuntze ) – senza caffeina

Per secoli, il tè verde è stato considerato una bevanda medicinale in Asia centrale e in Estremo Oriente. È ricco di polifenoli, un gruppo di sostanze chimiche dalle straordinarie capacità antiossidanti. Nel caso del tè verde, esiste un tipo di polifenoli chiamati catechine, responsabili del suo sapore genuino e del suo potere antiossidante. La principale catechina presente nel tè verde è l’epigallocatechina-3-gallato o EGCG. L’EGCG è 20 volte più antiossidante della vitamina E e 100 volte più della vitamina C [28]. In effetti, la maggior parte degli studi scientifici attribuisce gli effetti benefici del tè verde a livelli elevati di EGCG [28]. Questa sostanza protegge le cellule dai danni associati e, come nel caso della Bacopa, sopprime l’attività delle sostanze chimiche pro-infiammatorie prodotte nel corpo, come il fattore di necrosi tumorale alfa (TNF-α) [29]. Questa protezione può aiutare a ridurre i danni cerebrali che potrebbero portare al declino mentale e alle malattie del cervello come il Parkinson e l’Alzheimer. Studi basati sulla risonanza magnetica funzionale (MRI) hanno anche riscontrato benefici a breve termine nella memoria e nell’attenzione, nonché un aumento dell’attivazione di un’area del cervello responsabile della mediazione della memoria di lavoro [28, 29].
L’estratto di tè verde utilizzato nella formula MINFIRE è di prima qualità e utilizza un processo di estrazione avanzato che elimina la caffeina e trattiene i polifenoli antiossidanti (95% del suo contenuto totale). Dei polifenoli totali nel nostro estratto, l’80% sono catechine fenoliche e il 50% è costituito da EGCG puro. Per preparare 1 kg del nostro estratto di tè verde, è necessario utilizzare 100 kg di foglie di tè verde!
Colina L-bitartrato
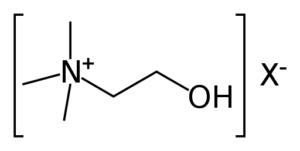
La colina non è una vitamina o un minerale, ma a causa delle sue somiglianze, è spesso raggruppata con le vitamine del complesso B. In effetti, questo nutriente influisce su una serie di funzioni corporee vitali come la funzionalità epatica, il movimento muscolare, il metabolismo, lo sviluppo sano del cervello, e la funzione del sistema nervoso [30].
Come accennato in precedenza, il sistema colinergico è modulato dal neurotrasmettitore acetilcolina, sintetizzato nell’organismo dalla colina e, quindi, ampiamente dipendente dall’assunzione giornaliera di questo nutriente [31]. Gli esseri umani possono produrre colina per via endogena nel fegato, principalmente come fosfatidilcolina, tuttavia la quantità che il corpo è in grado di produrre naturalmente non è sufficiente per soddisfare le esigenze quotidiane [32]. Molte persone, infatti, non raggiungono l’apporto consigliato di questo nutriente e possono soffrire di problemi di salute derivanti dalla sua carenza [15]. Esempi di alimenti che sono una fonte di colina: fegato, tuorli d’uovo, carne, pesce, broccoli e cavolfiore. Un’altra alternativa per superare un possibile deficit di colina è attraverso l’integrazione. L’integrazione con L-bitartrato di colina è una scelta eccellente, poiché ha un tasso di bioassorbimento molto più elevato rispetto alla colina semplice se assunta per via orale in capsule o compresse.
Gli effetti sulla memoria e sulla salute del cervello derivanti dall’integrazione di colina a lungo termine sono stati ampiamente studiati dalla comunità scientifica. Diversi studi in vivo su animali hanno osservato un aumento significativo delle concentrazioni di acetilcolina nel cervello dopo una dieta arricchita di colina [33]. Questi studi sono stati unanimi nell’attribuire un aumento significativo della memoria (memorizzazione della posizione del cibo) nei topi sottoposti a supplementazione di colina, sia nella fase prenatale [34-37], dopo la nascita [37, 38] o più tardi nella vita [39, 40]. Negli esseri umani, gli studi dimostrano che l’integrazione di colina può aiutare gli anziani a riprendersi da una moderata demenza di Alzheimer e da altri disturbi della memoria [41-44].
L-tirosina
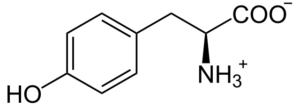
La L-tirosina è un amminoacido aromatico e un componente essenziale nella biosintesi di sostanze ad elevata attività fisiologica, come i neurotrasmettitori catecolamine dopamina, norepinefrina (norepinefrina) ed epinefrina (adrenalina) [45]. Infatti, studi scientifici hanno verificato una forte relazione tra l’assunzione di L-tirosina e una migliore sintesi di catecolamine nel cervello umano [46,47]. In questo senso, due studi clinici che confrontano pazienti con depressione e volontari sani hanno dimostrato che l’integrazione con L-tirosina ha migliorato positivamente il trattamento della depressione [48]. Il beneficio della supplementazione di L-tirosina è stato dimostrato anche in un altro studio clinico, in cui è stato chiesto a 22 adulti sani di passare rapidamente da un’attività all’altra. Rispetto al gruppo di controllo, la L-tirosina ha promosso positivamente la flessibilità cognitiva (una funzione cerebrale che si presume fosse regolata dalla dopamina) [49].
Un’altra importante funzione della tirosina è la produzione degli ormoni tiroidei tiroxina (T4) e triiodotironina (T3). Gli ormoni T3 e T4 regolano il metabolismo di proteine, grassi e carboidrati nel corpo e dirigono il modo in cui il corpo utilizza questi composti per produrre energia. La ghiandola tiroidea non può produrre questi due ormoni senza la presenza di L-tirosina e iodio. Pertanto, una carenza di entrambi può contribuire a bassi livelli di ormone tiroideo. Gli studi hanno scoperto che l’integrazione di L-tirosina può essere utile nel trattamento della stanchezza, un sintomo comune di bassi livelli di ormone tiroideo o surrenale [50].
La tirosina partecipa alla produzione di melanina, il pigmento responsabile del colore dei capelli e della pelle [51], aiuta nella funzione degli organi responsabili della produzione e della regolazione degli ormoni, come le ghiandole surrenali e ipofisarie (ipofisi), e partecipa al struttura di quasi tutte le proteine del corpo.
Fosfatidilserina
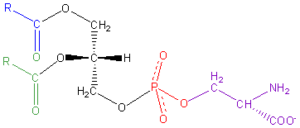
La fosfatidilserina è un fosfolipide di elevata qualità biologica, essenziale come nutriente per l’organismo. Sebbene sia distribuito in tutto il corpo, è nel cervello dove si accumula in maggiore quantità, soprattutto nella corteccia cerebrale o nella materia grigia [53]. La fosfatidilserina regola la conduzione degli impulsi nervosi, migliora la funzione e la memoria delle cellule nervose, partecipa al metabolismo dell’ATP e svolge un ruolo chiave nella segnalazione del ciclo cellulare, in particolare in relazione all’apoptosi [52, 53].
La concentrazione di fosfatidilserina nei nostri tessuti cerebrali diminuisce nel corso degli anni e questa diminuzione è correlata a una diminuzione delle capacità cognitive [52]. Uno studio clinico randomizzato, in doppio cieco e controllato con placebo ha concluso che l’integrazione di fosfatidilserina produce miglioramenti significativi nel disturbo da deficit di attenzione e iperattività (ADHD), nella memoria uditiva a breve termine, nella memoria di lavoro e nelle prestazioni mentali agli stimoli visivi. [54].
La fosfatidilserina utilizzata nei primi lavori scientifici per valutarne l’effetto benefico proveniva dal cervello bovino [55]. Tuttavia, a causa dei problemi di encefalopatia spongiforme bovina, il suo uso è stato sostituito dalla fosfatidilserina vegetale, ottenuta dalla lecitina di soia. Il contenuto naturale di fosfatidilserina nella lecitina di soia è di circa il 3%. Per aumentare questa quantità, viene eseguito un processo chiamato conversione enzimatica, che imita i processi enzimatici naturali. In MINFIRE viene utilizzata la fosfatidilserina di origine vegetale, derivata dalla lecitina di soia, con una concentrazione aumentata del 20%.
L’acido docosaesaenoico omega-3 (DHA) è il principale acido grasso polinsaturo nel sistema nervoso centrale e si accumula in particolare nella fosfatidilserina [52]. Per questo motivo, un corretto apporto di DHA è essenziale per la sintesi della fosfatidilserina nell’organismo [56]. Ciò è stato evidenziato in uno studio in vivo in cui è stato valutato l’effetto benefico sul corpo della combinazione di fosfatidilserina e DHA. Questo studio ha concluso che questa combinazione può ridurre significativamente la concentrazione di specie reattive dell’ossigeno nocive nel cervello e può migliorare le capacità di apprendimento e di memoria [57].
Il DHA è uno dei componenti di un altro prodotto esclusivo di Nutribiolite , OMEGA 3 + VIT K2 + VIT D3. Questo integratore alimentare contiene 1000 mg di acido grasso OMEGA 3 puro standardizzato al 50% di EPA (500 mg) e al 25% di DHA (250 mg). Raccomandiamo l’integrazione combinata di MINFIRE e OMEGA 3 + VIT K2 + VIT D3.