



Bacopa (Bacopa monnieri L.)
 Os efeitos benéficos da Bacopa (Bacopa monnieri L.) sobre a memória e a cognição foram extensivamente estudados e as suas propriedades nootrópicas se encontram descritas em vários artigos científicos de revisão [1-4]. A actividades benéficas da Bacopa para a saúde são atribuídas ao seu conteúdo de saponinas, principalmente dos bacósidos A e B [5]. Destas espécies químicas, os bacósidos A são os responsáveis por suas propriedades nootrópicas. De acordo com vários estudos, a Bacopa apresenta propriedades neuroprotectoras e de melhora das habilidades cognitivas, devido à sua capacidade de modular o sistema colinérgico e combater os processos responsáveis pelo stress oxidativo no cérebro [6-9]. O cérebro é especialmente susceptível ao stress oxidativo devido a sua alta actividade metabólica, seus níveis elevados de ferro, uma espécie susceptível a oxidação, e é composto por lipídios insaturados sujeitos à peroxidação [10].
Os efeitos benéficos da Bacopa (Bacopa monnieri L.) sobre a memória e a cognição foram extensivamente estudados e as suas propriedades nootrópicas se encontram descritas em vários artigos científicos de revisão [1-4]. A actividades benéficas da Bacopa para a saúde são atribuídas ao seu conteúdo de saponinas, principalmente dos bacósidos A e B [5]. Destas espécies químicas, os bacósidos A são os responsáveis por suas propriedades nootrópicas. De acordo com vários estudos, a Bacopa apresenta propriedades neuroprotectoras e de melhora das habilidades cognitivas, devido à sua capacidade de modular o sistema colinérgico e combater os processos responsáveis pelo stress oxidativo no cérebro [6-9]. O cérebro é especialmente susceptível ao stress oxidativo devido a sua alta actividade metabólica, seus níveis elevados de ferro, uma espécie susceptível a oxidação, e é composto por lipídios insaturados sujeitos à peroxidação [10].
O sistema colinérgico é formado por células nervosas que utilizam o neurotransmissor acetilcolina na transdução de sinais. Essas células produzem, liberam ou são activadas pela acetilcolina durante a propagação do impulso nervoso. O sistema colinérgico tem sido associado a várias funções cognitivas, como memória, atenção selectiva e processamento de emoções [11].
Outra propriedade importante da Bacopa é sua actividade anti-inflamatória. A neuroinflamação a longo prazo é prejudicial e pode conduzir ao surgimento de doenças neurodegenerativas como Alzheimer, Parkinson e esclerose múltipla [12]. Estudos in vitro com células microgliais (células do tecido nervoso) demonstraram que os extractos alcalóides de Bacopa, assim como o bacosídeo A, inibem a liberação de citocinas pró-inflamatórias, como o Factor de necrose tumoral alfa (TNF-α) e interleucina 6 (IL-6) [13]. Esses compostos também foram relacionados com a inibição de enzimas associadas à inflamação no cérebro [12].
O extracto de Bacopa monnieri de MINFIRE é obtido a partir de métodos de extracção avançados, nos quais se obtém um extracto com um alto grau de pureza e alta concentração de bacósidos. Para produzir apenas 1 kg do nosso extracto e atingir sua concentração de 50% em bacósidos, são necessários 30 kg de Bacopa! A concentração de 50% de bacósidos em nosso extracto é uma das mais altas do mercado, já que normalmente os extractos usados em outros produtos são da ordem de 25% -45%. Além disso, nosso extracto atende a todos os requisitos de qualidade da indústria de suplementos alimentares e aos critérios exigentes dos produtos Nutribiolite.
Hericium ou Juba de leão (Hericium erinaceus Bull. Pers.)
 O Hericium, também conhecido como juba de leão, é um fungo que se distribui entre a América do Norte e a Ásia. Este fungo tem uma longa história na medicina tradicional chinesa, sendo-lhe atribuídas propriedades benéficas para a saúde do cérebro [14]. De fato, estudos mais recentes atribuem grande potencial ao Hericium no tratamento de doenças neurológicas, uma vez que contém compostos neurotróficos bioativos que podem atravessar a barreira hematoencefálica [15, 16]. Este fungo é abundante em compostos bioativos como os polissacáridos β-glucanos e os terpenóides hericenona e erinacina.
O Hericium, também conhecido como juba de leão, é um fungo que se distribui entre a América do Norte e a Ásia. Este fungo tem uma longa história na medicina tradicional chinesa, sendo-lhe atribuídas propriedades benéficas para a saúde do cérebro [14]. De fato, estudos mais recentes atribuem grande potencial ao Hericium no tratamento de doenças neurológicas, uma vez que contém compostos neurotróficos bioativos que podem atravessar a barreira hematoencefálica [15, 16]. Este fungo é abundante em compostos bioativos como os polissacáridos β-glucanos e os terpenóides hericenona e erinacina.
Os β -glucanos têm sido alvo de particular interesse devido à sua bioatividade. Por exemplo, estudos in vivo identificaram efeitos protetores destes na hepatotoxicidade oxidativa em ratos [17] Os efeitos neuroprotectores dos β-glucanos também foram observados em estudos in vitro com células tóxicas pela formação de placas β-amilóide. As placas β-amilóides são depósitos de proteínas que normalmente ocorrem no cérebro e estão associados à doença de Alzheimer, asfixiando as células cerebrais e contribuindo para a degeneração progressiva dos neurónios. Nesses estudos, os β-glucanos promoveram a viabilidade celular e protegeram as células contra a apoptose induzida pela formação de placas β-amilóide [18].
A erinacina é uma família de moléculas orgânicas denominadas terpenóides, capazes de atravessar a barreira hematoencefálica e atuar diretamente no cérebro. O grande interesse pelos extratos de Hericium é devido principalmente à presença de erinacina. Vários estudos científicos têm demostrado que esta molécula é capaz de induzir a síntese da proteína Fator de Crescimento Nervoso (NGF) [16, 19], essencial para o desenvolvimento e manutenção dos neurónios do sistema nervoso periférico e para integridade funcional dos neurónios colinérgicos (do sistema colinérgico) presentes no sistema nervoso central [20].
Em MINFIRE, o fungo Hericium ou juba de leão passa por um avançado processo de extração com uma mistura hidroalcoólica de etanol/água para obter uma maior quantidade de moléculas orgânicas solúveis como a erinacina. Para produzir apenas 1kg do nosso extrato de juba de leão, são necessários 10 kg de cogumelo. Esses extratos são normalmente padronizados em termos de polissacáridos totais. Quanto menos polissacárido, mais compostos orgânicos como a erinacina estarão presentes. Por este motivo, selecionamos para MINFIRE um extrato com 30% de polissacarídeos. Segundo nossa experiência, este extrato apresenta a melhor relação entre β-glucanos e erinacina. Outros extratos com maior proporção em polissacáridos, apresentarão uma menor quantidade de erinacina.
Ginkgo (Ginkgo biloba L.)
 Com o avanço da idade, o corpo experimenta uma diminuição da elasticidade e tonicidade dos vasos sanguíneos, o que limita a fluidez do sangue. A insuficiência de sangue, oxigénio e nutrientes no cérebro pode prejudicar a função cognitiva. O extracto de Ginkgo biloba exerce efeitos dilatadores e constritivos nos vasos sanguíneos, aumentando o fluxo sanguíneo para o cérebro e, consequentemente, seus níveis de glicose e ATP, resultando numa melhora da função mental [21]. Estudos in vivo demonstraram que os compostos bioactivos do Ginkgo, os glucósidos flavonóides e as lactonas de terpeno, atravessam facilmente a barreira hematoencefálica, agindo directamente no cérebro [22]. Estes compostos actuam através da actividade antioxidante e antiplaquetária (PAF), dos efeitos moduladores sobre o tónus cerebrovascular, da actividade de receptor/transmissor, do metabolismo de glicose e da actividade electroencefalográfica [22-26]. Como no caso do extracto de Bacopa, estudos recentes sugeriram que o extracto de Ginkgo também tem efeitos directos no sistema colinérgico, aumentando a capacidade da acetilcolina de se ligar a um local do receptor sináptico, o que poderia explicar seus efeitos estimulantes cognitivos tanto agudos quanto crónicos [27].
Com o avanço da idade, o corpo experimenta uma diminuição da elasticidade e tonicidade dos vasos sanguíneos, o que limita a fluidez do sangue. A insuficiência de sangue, oxigénio e nutrientes no cérebro pode prejudicar a função cognitiva. O extracto de Ginkgo biloba exerce efeitos dilatadores e constritivos nos vasos sanguíneos, aumentando o fluxo sanguíneo para o cérebro e, consequentemente, seus níveis de glicose e ATP, resultando numa melhora da função mental [21]. Estudos in vivo demonstraram que os compostos bioactivos do Ginkgo, os glucósidos flavonóides e as lactonas de terpeno, atravessam facilmente a barreira hematoencefálica, agindo directamente no cérebro [22]. Estes compostos actuam através da actividade antioxidante e antiplaquetária (PAF), dos efeitos moduladores sobre o tónus cerebrovascular, da actividade de receptor/transmissor, do metabolismo de glicose e da actividade electroencefalográfica [22-26]. Como no caso do extracto de Bacopa, estudos recentes sugeriram que o extracto de Ginkgo também tem efeitos directos no sistema colinérgico, aumentando a capacidade da acetilcolina de se ligar a um local do receptor sináptico, o que poderia explicar seus efeitos estimulantes cognitivos tanto agudos quanto crónicos [27].
O extracto padronizado de Ginkgo de MINFIRE contém no mínimo 24% de glicosídeos flavonóides e 6% de lactonas de terpeno. É um dos extractos mais valorizados e utilizados na prevenção da demência e deterioração cognitiva. Para preparar apenas 1kg de nosso extracto concentrado, é necessário utilizar 50 kg de folha seca de Ginkgo!
Chá verde (Camellia sinensis L. Kuntze ) – sem cafeína
 Durante séculos, o chá verde foi considerado uma bebida medicinal na Ásia Central e no Extremo Oriente. É rico em polifenóis, um grupo de produtos químicos com extraordinária capacidade antioxidante. No caso do chá verde, existe um tipo de polifenóis denominados catequinas, responsáveis pelo seu sabor genuíno e pelo seu poder antioxidante. A principal catequina presente no chá verde é a epigalocatequina-3-galato ou EGCG. O EGCG é 20 vezes mais antioxidante do que a vitamina E, e 100 vezes mais antioxidante do que a vitamina C [28]. Com efeito, a maioria dos estudos científicos atribuem os efeitos benéficos do chá verde aos altos níveis de EGCG [28]. Esta substância protege as células do stress oxidativo e suprime a actividade de produtos químicos pró-inflamatórios produzidos no organismo, como o factor de necrose tumoral alfa (TNF-α) [29]. Esta protecção pode ajudar a reduzir os danos cerebrais que podem conduzir ao declínio mental e a doenças cerebrais como Parkinson e Alzheimer. Estudos baseados em ressonância magnética funcional (MRI) também encontraram benefícios de curto prazo na memória e na atenção, bem como um aumento na activação de uma área do cérebro responsável por mediar a memória de trabalho [28, 29].
Durante séculos, o chá verde foi considerado uma bebida medicinal na Ásia Central e no Extremo Oriente. É rico em polifenóis, um grupo de produtos químicos com extraordinária capacidade antioxidante. No caso do chá verde, existe um tipo de polifenóis denominados catequinas, responsáveis pelo seu sabor genuíno e pelo seu poder antioxidante. A principal catequina presente no chá verde é a epigalocatequina-3-galato ou EGCG. O EGCG é 20 vezes mais antioxidante do que a vitamina E, e 100 vezes mais antioxidante do que a vitamina C [28]. Com efeito, a maioria dos estudos científicos atribuem os efeitos benéficos do chá verde aos altos níveis de EGCG [28]. Esta substância protege as células do stress oxidativo e suprime a actividade de produtos químicos pró-inflamatórios produzidos no organismo, como o factor de necrose tumoral alfa (TNF-α) [29]. Esta protecção pode ajudar a reduzir os danos cerebrais que podem conduzir ao declínio mental e a doenças cerebrais como Parkinson e Alzheimer. Estudos baseados em ressonância magnética funcional (MRI) também encontraram benefícios de curto prazo na memória e na atenção, bem como um aumento na activação de uma área do cérebro responsável por mediar a memória de trabalho [28, 29].
O extracto de chá verde da fórmula de MINFIRE é de qualidade Premium e usa um processo de extracção avançado que elimina a cafeína à medida que aumenta sua concentração em polifenóis antioxidantes (95% de seu conteúdo total). Do total de polifenóis em nosso extracto, 80% são catequinas fenólicas e 50% consistem em EGCG puro. Para preparar 1 kg do nosso extracto de chá verde, é necessário utilizar 100 kg de folhas de chá verde!
L-bitartarato de colina
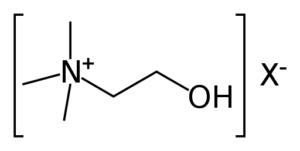
A colina apesar de não ser uma vitamina é frequentemente agrupada com as vitaminas do complexo B. Na verdade, esse nutriente afecta uma série de funções corporais vitais, como função hepática, movimento muscular, metabolismo, desenvolvimento saudável do cérebro e funcionamento do sistema nervoso [30].
Como mencionado anteriormente, o sistema colinérgico é modulado pelo neurotransmissor acetilcolina, sintetizado no organismo a partir da colina e, portanto, amplamente dependente da ingestão diária desse nutriente [31]. Os seres humanos podem produzir colina endogenamente no fígado, principalmente como fosfatidilcolina, porém a quantidade que o corpo é capaz de produzir naturalmente não é suficiente para atender às necessidades diárias [32]. Com efeito, muitas pessoas não atingem a ingestão recomendada desse nutriente e podem sofrer problemas de saúde decorrentes de sua deficiência [31]. Exemplos de alimentos que são fonte de colina: fígado, gema de ovo, carne, peixe, brócolos e couve-flor. Outra alternativa para superar um possível deficit de colina é suplementação. Para isso o L-bitartarato de colina é uma escolha excelente, pois, devido a sua maior solubilidade, apresenta uma taxa de bioabsorção muito mais alta do que a colina simples, quando tomada por via oral em cápsulas ou comprimidos.
Os efeitos benéficos na memória e na saúde do cérebro, derivados da suplementação continuada com colina, foram amplamente estudados pela comunidade científica. Vários estudos in vivo em animais verificaram um aumento significativo nas concentrações de acetilcolina no cérebro após uma dieta enriquecida com colina [33]. Esses estudos foram unânimes em atribuir um aumento significativo na memória (memorização da localização do alimento) em ratos submetidos à suplementação de colina, tanto na fase pré-natal [34-37], após o nascimento [37, 38] ou na vida adulta [39, 40]. Estudos em humanos, concluíram que a suplementação com colina pode ajudar os idosos a se recuperarem da demência moderada de Alzheimer e de outras alterações da memória [41-44].
L-tirosina
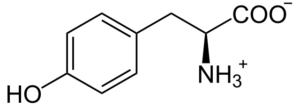 A tirosina é um aminoácido aromático e um componente essencial na biossíntese de substâncias com alta actividade fisiológica, como os neurotransmissores dopamina, norepinefrina (noradrenalina) e epinefrina (adrenalina) [45]. Na verdade, estudos científicos verificaram uma forte relação entre a ingestão de L-tirosina e a melhora da síntese de catecolaminas no cérebro em humanos [46, 47]. A este respeito, dois estudos clínicos comparando pacientes com depressão e voluntários saudáveis mostraram que a suplementação com L-tirosina melhorou positivamente o tratamento da depressão [48]. O benefício da suplementação de L-tirosina também foi demonstrado em outro estudo no qual 22 adultos saudáveis foram solicitados a alternar rapidamente entre duas tarefas diferentes. Em comparação com o grupo de controle, a L-tirosina promoveu positivamente a flexibilidade cognitiva (uma função cerebral que se presume ser regulada pela dopamina) [49].
A tirosina é um aminoácido aromático e um componente essencial na biossíntese de substâncias com alta actividade fisiológica, como os neurotransmissores dopamina, norepinefrina (noradrenalina) e epinefrina (adrenalina) [45]. Na verdade, estudos científicos verificaram uma forte relação entre a ingestão de L-tirosina e a melhora da síntese de catecolaminas no cérebro em humanos [46, 47]. A este respeito, dois estudos clínicos comparando pacientes com depressão e voluntários saudáveis mostraram que a suplementação com L-tirosina melhorou positivamente o tratamento da depressão [48]. O benefício da suplementação de L-tirosina também foi demonstrado em outro estudo no qual 22 adultos saudáveis foram solicitados a alternar rapidamente entre duas tarefas diferentes. Em comparação com o grupo de controle, a L-tirosina promoveu positivamente a flexibilidade cognitiva (uma função cerebral que se presume ser regulada pela dopamina) [49].
Outra função importante da tirosina é a produção dos hormônios da glândula tiróide a tiroxina (T4) e triiodotironina (T3). Os hormônios T3 e T4 são responsáveis por regular o metabolismo das proteínas, gorduras e carboidratos. A glândula tiróide não pode produzir esses hormônios sem a presença da tirosina e do iodo. Portanto, a deficiência de ambos nutrientes pode conduzir a sérios problemas hormonais. Com efeito, a suplementação com L-tirosina pode ser benéfica no tratamento da fadiga, um sintoma muito comum quando se sofre de baixos níveis de hormônios tireoidianos ou adrenais [50].
A tirosina participa também na produção de melanina, o pigmento responsável pela cor do cabelo e da pele [51], auxilia no funcionamento dos órgãos responsáveis pela produção e regulação de hormônios, como as glândulas adrenais e pituitária (hipófise), e está envolvida na a estrutura de quase todas as proteínas do corpo.
Fosfatidilserina
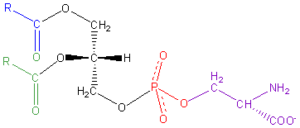 A fosfatidilserina é um fosfolípido, uma gordura de elevada qualidade biológica, essencial como nutriente para o organismo. Embora esteja distribuída por todo o corpo, é no cérebro onde se acumula em maior quantidade, especialmente no córtex cerebral ou substância cinzenta [52]. A fosfatidilserina regula a condução dos impulsos nervosos, melhora a função e a memória das células nervosas, participa do metabolismo do ATP e desempenha um papel fundamental na sinalização do ciclo celular, especificamente em relação à apoptose [53, 54].
A fosfatidilserina é um fosfolípido, uma gordura de elevada qualidade biológica, essencial como nutriente para o organismo. Embora esteja distribuída por todo o corpo, é no cérebro onde se acumula em maior quantidade, especialmente no córtex cerebral ou substância cinzenta [52]. A fosfatidilserina regula a condução dos impulsos nervosos, melhora a função e a memória das células nervosas, participa do metabolismo do ATP e desempenha um papel fundamental na sinalização do ciclo celular, especificamente em relação à apoptose [53, 54].
A concentração de fosfatidilserina nos tecidos cerebrais diminui com o passar dos anos, e está relacionado com a diminuição nas habilidades cognitivas [53]. Um ensaio clínico randomizado, duplo-cego e controlado por placebo concluiu que a suplementação de fosfatidilserina produz melhorias significativas no transtorno de deficit de atenção e hiperactividade (TDAH), memória auditiva de curto prazo, memória de trabalho e desempenho mental para estímulos visuais [55].
O ácido docosahexaenóico (DHA) do Omega 3 é o principal ácido graxo polinsaturado do sistema nervoso central e se armazena particularmente na fosfatidilserina [53]. Por este motivo, um fornecimento correcto de DHA é essencial para a biossíntese da fosfatidilserina [56]. De facto, a combinação de fosfatidilserina e DHA pode reduzir significativamente a concentração de radicais livres no cérebro e melhorar as capacidades de aprendizagem e memória [57].
O DHA é um dos componentes de outro produto incrível da Nutribiolite, o OMEGA 3 + VIT K2 + VIT D3. Este suplemento alimentar tem 1000 mg de ácido graxo OMEGA 3 puro padronizado para 50% de EPA (500 mg) e 25% em DHA (250 mg).
A fosfatidilserina utilizada nos primeiros trabalhos científicos, tinha como origem o cérebro bovino [58]. No entanto, devido aos problemas relacionados com a encefalopatia espongiforme bovina, o seu uso foi paulatinamente substituído pela fosfatidilserina de origem vegetal, obtida a partir da lecitina de soja. O conteúdo natural de fosfatidilserina na lecitina de soja é de apenas 3%. Para aumentar essa quantidade, é realizado um processo denominado conversão enzimática, que simula processos enzimáticos naturais. MINFIRE utiliza fosfatidilserina de origem vegetal, derivada da lecitina de soja, e concentrada por conversão enzimática até alcançar a concentração de 20%.