



Bacopa (Bacopa monnieri L.)
 Los efectos benéficos de la bacopa (Bacopa monnieri L.) sobre la memoria y la cognición han sido extensivamente estudiados y muchos artículos científicos de revisión han descrito sus propiedades nootrópicas [1-4]. La actividad de la bacopa ha sido atribuida a su contenido de saponinas, principalmente los bacósidos A y B [5]. De estas especies química, la que se conoce responsable por sus propiedades nootrópicas es el bacósido A. Según varios estudios, se ha sugerido que la bacopa posé propiedades neuroprotectoras y de mejora de las capacidades cognitivas, debido a su capacidad de modular el sistema colinérgico y combatir los procesos responsable por el daño oxidativo en el cerebro [6-9]. El cerebro es especialmente susceptible de sufrir al daño oxidativo, debido a su elevada actividad metabólica, a sus niveles elevados de hierro, que es una especie prooxidante (que induce el daño oxidativo) y está compuestos por lípidos insaturados, propensos a la peroxidación [10].
Los efectos benéficos de la bacopa (Bacopa monnieri L.) sobre la memoria y la cognición han sido extensivamente estudiados y muchos artículos científicos de revisión han descrito sus propiedades nootrópicas [1-4]. La actividad de la bacopa ha sido atribuida a su contenido de saponinas, principalmente los bacósidos A y B [5]. De estas especies química, la que se conoce responsable por sus propiedades nootrópicas es el bacósido A. Según varios estudios, se ha sugerido que la bacopa posé propiedades neuroprotectoras y de mejora de las capacidades cognitivas, debido a su capacidad de modular el sistema colinérgico y combatir los procesos responsable por el daño oxidativo en el cerebro [6-9]. El cerebro es especialmente susceptible de sufrir al daño oxidativo, debido a su elevada actividad metabólica, a sus niveles elevados de hierro, que es una especie prooxidante (que induce el daño oxidativo) y está compuestos por lípidos insaturados, propensos a la peroxidación [10].
El sistema colinérgico: Está compuesto por células nerviosas organizadas que utilizan el neurotransmisor acetilcolina en la transducción de señales. Estas células producen, liberan o son activadas por la acetilcolina durante la propagación del impulso nervioso. El sistema colinérgico ha sido asociado a varias funciones cognitivas como la memoria, la atención selectiva y el procesamiento de las emociones [11].
Otra importante propiedad de la bacopa es su actividad antiinflamatoria en el cerebro [12]. La neuroinflamación, a largo plazo es perjudicial y puede provocar neurodegeneración, que a largo plazo, puede conducir al surgimiento de enfermedades neurodegenerativas como el Alzheimer, Parkinson, y esclerosis múltiple [12]. Estudios in vitro con células microgliales (células del tejido nervioso) han demostrado que los extractos alcaloides de la bacopa, bien como el bacósido A, inhiben la liberación de las citoquinas proinflamatorias como el factor de necrosis tumoral alfa (TNF-α) y la interleuquina 6 (IL-6) [13]. Estos compuestos fueron también relacionados con la inhibición de enzimas asociadas a la inflamación en el cerebro [12].
El extracto de Bacopa monnieri de MINFIRE se obtiene a partir de métodos de extracción avanzados, en el cual se obtiene un extracto con un elevado grado de pureza y una elevada concentración en bacósidos. ¡Para producir solamente 1 kg de nuestro extracto 30:1 y alcanzar su concentración mínima de 50% en bacósidos, son necesarios 30 kg de bacopa! La concentración de 50% en bacósidos de nuestro extracto es de la más elevada del mercado, una vez que normalmente los extractos que se utilizan en otros productos son del orden del 25%-45% en bacósidos. Además, nuestro extracto cumple con todos los requisitos de calidad de la industria de complementos alimenticios y los criterios muy exigentes de los productos Nutribiolite.
Hericium o Mellena de león (Hericium erinaceus Bull. Pers.)
 El Hericium también conocido como melena de león, es un hongo que se encuentra distribuido entre América del Norte y Asia. Este hongo tiene una larga historia en la medicina tradicional china, siéndole atribuido propiedades benéficas para la salud del cerebro [14]. De hecho, estudios más recientes atribuyen al hericium un grande potencial en el tratamiento de enfermedades neurológicas, una vez que contiene compuestos neurotróficos bioactivos que puedes pasar a través de la barrera hematoencefálica [15, 16]. Este hongo es abundante en compuestos bioactivos como los polisacáridos β-glucanos y los terpenoides hericenona y erinacina.
El Hericium también conocido como melena de león, es un hongo que se encuentra distribuido entre América del Norte y Asia. Este hongo tiene una larga historia en la medicina tradicional china, siéndole atribuido propiedades benéficas para la salud del cerebro [14]. De hecho, estudios más recientes atribuyen al hericium un grande potencial en el tratamiento de enfermedades neurológicas, una vez que contiene compuestos neurotróficos bioactivos que puedes pasar a través de la barrera hematoencefálica [15, 16]. Este hongo es abundante en compuestos bioactivos como los polisacáridos β-glucanos y los terpenoides hericenona y erinacina.
Los β-glucanos han sido objeto de particular interés debido su bioactividad. Por ejemplo, estudios in vivo han identificado efectos protectores de estos sobre la hepatotoxicidad oxidativa en ratones [17]. Se observaron, también, efectos neuroprotectores de los β-glucano en estudios in vitro con células tóxicas por la formación de placas β-amiloide. Las placas β-amiloide son depósitos de proteínas, que normalmente ocurren en el cerebro y están asociadas a la enfermedad de Alzheimer, asfixiando las células del cerebro y contribuyendo a la degeneración progresiva de las neuronas. Los polisacáridos de β-glucano también promovieron la viabilidad celular y protegieron a las células contra la apoptosis inducida por la formación de placas β-amiloide [18].
La erinacina es una familia de moléculas orgánicas, denominadas terpenoides, capaces de atravesar la barrera hematoencefálica y actuar directamente en el cerebro. El gran interés en los extractos de Hericium se basa principalmente en la presencia de la erinacina. Se ha demostrado en varios estudios científicos que esta molécula es capaz de inducir la síntesis de la proteína Factor de crecimiento nerviosos (NGF) [16, 19], esencial para el desarrollo y mantenimiento de las neuronas del sistema nervioso periférico y para la integridad funcional de las neuronas colinérgicas (del sistema colinérgico) presentes en el sistema nervioso central [20].
En MINFIRE, el hongo Hericium erinaceus o melena de león se somete a un proceso de extracción avanzada con una mezcla hidroalcohólica de etanol:agua para obtener una mayor cantidad de moléculas orgánicas solubles como la erinacina. Para producir nuestro extracto de melena de león, son necesarios 10 kg del hongo para producir solamente 1kg de extracto. Estos extractos son normalmente estandarizados en términos de polisacáridos totales. Cuanto menos polisacárido, más compuestos orgánicos como la erinacina estarán presentes, por este motivo nuestro extracto contiene un 30% de polisacáridos, para tener la mejor relación entre polisacáridos β-glucanos y erinacina. Otros extractos presentan más polisacáridos lo que va disminuir su contenido en erinacina.
Ginkgo (Ginkgo biloba L.)

El extracto de las hojas del ginkgo sigue siendo uno de los extractos de plantas más populares utilizados en Europa para aliviar los síntomas asociados con una variedad de trastornos cognitivos como la enfermedad de Alzheimer, la demencia vascular y las afecciones amnésicas relacionadas con la edad [21]. Se ha demostrado en estudios científicos con sujetos tanto de edad joven como de edad avanzada, el efecto benéfico de la suplementación con el extracto de ginkgo sobre la memoria a corto [22, 23] y a largo plazo [24, 25]. También se han demostrado el efecto positivo de la suplementación con ginkgo en la mejora de la memoria en pacientes ancianos diagnosticados con enfermedad cerebrovascular [26] y un efecto modesto, pero significativo, en pacientes con enfermedad de Alzheimer [27].
El conocimiento exacto de mecanismo de acción de extracto de ginkgo aún es incierto, debido a la complexidad de los principios activos que lo constituyen, pero se sabe que los efectos neuroprotectores y de mejora cognitiva son debidos, en gran medida, a la presencia de glucósidos flavonoides y de lactonas de terpeno (también conocidas como ginkgólidos) [28]. Los glucósidos flavonoides son sobre todo potentes antioxidantes y neutralizadores de radicales libres y las lactonas terpénicas, en particular el ginkgólido B, son potentes antagonistas del factor activador de plaquetas, también conocido como PAF (platelet-activating factor, por sus siglas en inglés). El PAF es un es un potente activador de fosfolípidos que interviene como mediador en muchas funciones de los leucocitos, incluyendo agregación de plaquetas, procesos isquémicos, inflamación y reacciones de tipo anafiláctico [29]. Se ha demostrado que los ginkgólidos bloquean los receptores de PAF de las células blanco, previniendo la ocurrencia de estos procesos inflamatorios como el aumento de la permeabilidad vascular [30] y efectos directos sobre la función neuronal [31, 32]. Los otros componentes principales del extracto de ginkgo son los glucósidos flavonoides, que contribuyen a los efectos antioxidantes y de neutralizador de radicales libres [33]. Existe una creciente evidencia de que el estrés oxidativo juega un papel importante en el proceso normal de envejecimiento cerebral y en la fisiopatología de trastornos neurodegenerativos como la enfermedad de Alzheimer [34-37]. Estudios recientes han demostrado que el extracto de ginkgo ejerce efectos neuroprotectores en las neuronas del cerebro contra el estrés oxidativo inducido por la peroxidación lipídica [38-40]. El estrés oxidativo ha sido estudiado fundamentalmente en las enfermedades neurodegenerativas como la enfermedad de Alzheimer, la enfermedad de Parkinson y la esclerosis lateral amiotrófica [41]. En estas enfermedades se ha encontrado daño oxidativo incluso en etapas tempranas de la enfermedad, lo que indica que los radicales libres están relacionados con la etiología de estas.
Si bien los efectos de mejora cognitiva del ginkgo se han atribuido, principalmente a sus efectos antagonistas del PAF y neutralizadora de los radicales libres, estudios recientes en animales sugieren que los componentes del extracto de Ginkgo pueden tener efectos directos sobre el sistema colinérgico, lo que podría explicar sus efectos de mejora cognitiva tanto agudos (que se producen durante o inmediatamente después) como crónicos (efectos producidos por la toma a largo plazo) [42].
Las acciones colinérgicas directas del ginkgo observadas en estudios en animales incluyen la reducción de la amnesia inducida por administración de escopolamina [43], la modulación de la captación de colina presináptica en las terminales de las neuronas y la liberación de acetilcolina [44], la regulación al alza de los receptores muscarnicos postsinápticos y los efectos indirectos sobre la función colinérgica por modulación del sistema serotoninérgico [45].
El extracto de hoja de Ginkgo biloba de MINFIRE está concentrado 50 veces en relación al polvo de hoja seca y titulado en 24% en glucósidos flavonoides y 6% en lactonas de terpeno por HPLC (cromatografía liquida de alta performance).
Té verde (Camellia sinensis L. Kuntze ) – sin cafeína

Por siglos, el té verde ha sido considerado una bebida medicinal en Asia central y lejano Oriente. Es rico en polifenoles, un grupo de sustancias química con una extraordinaria capacidad antioxidante. En el caso del té verde, existe un tipo de polifenoles denominados catequinas, responsables de su sabor genuino y por su poder antioxidante. La principal catequina presente en el té verde es la epigalocatequina-3-galato o EGCG. La EGCG es 20 veces más antioxidante que la vitamina E y 100 veces más que la vitamina C [46]. De hecho, la mayoría de los estudios científicos atribuyen los efectos beneficiosos del té verde a los altos niveles de EGCG. Esta sustancia protege las células del daño asociado al estrés oxidativo y suprime la actividad de las sustancias químicas proinflamatorias producidas en el cuerpo, como el factor de necrosis tumoral alfa (TNF-α) [47]. Esta protección puede ayudar a reducir el daño cerebral que podría conducir al deterioro mental y enfermedades cerebrales como el Parkinson y el Alzheimer. Estudios basados en imágenes de resonancia magnética funcional (MRI) también encontraron beneficios a corto plazo en la memoria y la atención, así como un aumento en la activación de un área del cerebro responsable de mediar la memoria de trabajo [46, 47].
El extracto de té verde utilizado en la fórmula de MINFIRE es de calidad Premium y utiliza un proceso avanzado de extracción que retiene los polifenoles antioxidantes (95% de su contenido total). De los polifenoles totales de nuestro extracto, 80% son catequinas fenólicas y 50% consiste en EGCG puro. ¡Para preparar 1 kg de nuestro extracto de té verde, es necesario utilizar 100 kg de hojas de té verde!
L-bitartrato de colina
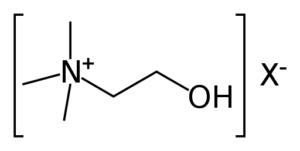 La colina no es una vitamina ni un mineral, pero, debido a sus similitudes, a menudo se agrupa con las vitaminas del complejo B. De hecho, este nutriente afecta una serie de funciones corporales vitales como la función hepática, el movimiento muscular, el metabolismo, el desarrollo saludable del cerebro y el funcionamiento del sistema nervioso [48].
La colina no es una vitamina ni un mineral, pero, debido a sus similitudes, a menudo se agrupa con las vitaminas del complejo B. De hecho, este nutriente afecta una serie de funciones corporales vitales como la función hepática, el movimiento muscular, el metabolismo, el desarrollo saludable del cerebro y el funcionamiento del sistema nervioso [48].
Como mencionado anteriormente, el sistema colinérgico está modulado por el neurotransmisor acetilcolina, sintetizado en el cuerpo a partir de la colina y, por eso, dependiente en gran medida de la ingesta diaria de este nutriente [49]. Los seres humanos pueden producir colina de forma endógena en el hígado, principalmente como fosfatidilcolina, sin embargo, la cantidad que el cuerpo es capaz de producir de forma natural no es suficiente para satisfacer las necesidades humanas diarias [50]. De hecho, muchas personas no alcanzan la ingesta recomendada de este nutriente y pueden sufrir de los problemas de salud derivados de su deficiencia [15]. Ejemplos de alimentos que son fuente de colina: hígado, yemas de huevo, carne, pescado, brócoli y coliflor. Otra alternativa para superar un eventual déficit en colina es a través de la suplementación. La suplementación con L-bitartrato de colina es una excelente elección, una vez que presenta una tasa de bioabsorción muy superior a la colina simple a través de la toma oral como cápsulas o comprimidos.
Los efectos sobre la memoria y la salud del cerebro derivados de la suplementación a largo plazo con colina, han sido ampliamente estudiados por la comunidad científica. Varios estudios in vivo en animales han observado un incremento importante en las concentraciones de acetilcolina en el cerebro después de una dieta enriquecida con colina [51]. Estos estudios han sido unánimes en atribuir un incremento importante en la memoria (memorización de la ubicación de los alimentos) en ratones objeto de suplementación con colina, tanto en la fase prenatal [52-55], posteriormente al nacimiento [55, 56] o más tarde en vida [57, 58]. En humanos, los estudios muestran que la suplementación con colina puede ayudar a los ancianos a recuperarse de la demencia moderada de Alzheimer y otras alteraciones de la memoria [59-62].
L-tirosina
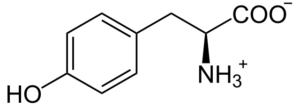 La L-tirosina es un aminoácido aromático y componente esencial en la biosíntesis de substancias con alta actividad fisiológica, como las catecolaminas neurotransmisores dopamina, norepinefrina (noradrenalina) y epinefrina (adrenalina) [63]. De hecho, estudios científicos han verificado una fuerte relación entre la ingesta de L-tirosina y la mejora de la síntesis de catecolaminas en el cerebro en seres humanos [64, 65]. En relación con eso, dos estudios clínicos comparando pacientes con depresión y voluntarios sanos han demostrado que la suplementación con L-tirosina mejoró de forma positiva el manejo de la depresión [66]. El beneficio de la suplementación con L-tirosina también ha quedado demostrado en otro estudio clínico, en el cual se pidió a 22 adultos sanos que cambiaran rápidamente entre dos tareas diferentes. En comparación con el grupo de control, la L-tirosina promovió positivamente la flexibilidad cognitiva (una función cerebral que se supone está regulada por la dopamina) [67].
La L-tirosina es un aminoácido aromático y componente esencial en la biosíntesis de substancias con alta actividad fisiológica, como las catecolaminas neurotransmisores dopamina, norepinefrina (noradrenalina) y epinefrina (adrenalina) [63]. De hecho, estudios científicos han verificado una fuerte relación entre la ingesta de L-tirosina y la mejora de la síntesis de catecolaminas en el cerebro en seres humanos [64, 65]. En relación con eso, dos estudios clínicos comparando pacientes con depresión y voluntarios sanos han demostrado que la suplementación con L-tirosina mejoró de forma positiva el manejo de la depresión [66]. El beneficio de la suplementación con L-tirosina también ha quedado demostrado en otro estudio clínico, en el cual se pidió a 22 adultos sanos que cambiaran rápidamente entre dos tareas diferentes. En comparación con el grupo de control, la L-tirosina promovió positivamente la flexibilidad cognitiva (una función cerebral que se supone está regulada por la dopamina) [67].
Otra función importante de la tirosina es la producción de hormonas tiroideas tiroxina (T4) y triyodotironina (T3). Las hormonas T3 y T4 regulan el metabolismo de proteínas, grasas y carbohidratos del cuerpo, y dirigen cómo el cuerpo usa estos compuestos para producir energía. La glándula tiroides no puede producir estas dos hormonas sin la presencia del L-tirosina y el yodo. Por lo tanto, una deficiencia de ambos puede contribuir a niveles bajos de hormona tiroidea. Los estudios han verificado que la suplementación con L-tirosina puede ser beneficiosa para tratar la fatiga, un síntoma común de niveles bajos de hormonas tiroideas o suprarrenales [68].
La tirosina participa en la producción de la melanina, el pigmento responsable del color del cabello y la piel [69], ayuda en la función de los órganos responsables de producir y regular las hormonas, como las glándulas suprarrenales y pituitaria (hipófisis) y, está involucrado en la estructura de casi todas las proteínas del cuerpo.
Fosfatidilserina
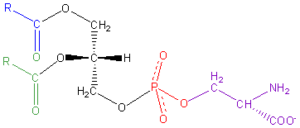
La fosfatidilserina es un fosfolípido componente fundamental de la membrana celular. Aunque se distribuye por todo el organismo, es en el cerebro donde se acumula en mayor cantidad, especialmente en la corteza cerebral o materia gris [70]. La fosfatidilserina regula la conducción de los impulsos nerviosos, mejora la función y la memoria de las células nerviosas, participa en el metabolismo del adenosín trifosfato (ATP) y desempeña un papel clave en la señalización del ciclo celular, específicamente en relación con la apoptosis [71, 72].
La concentración de fosfatidilserina en nuestros tejidos cerebrales disminuye con el paso de los años y esta disminución está relacionada con una disminución de las capacidades cognitivas [71]. Un ensayo clínico aleatorizado, doble ciego y controlado con placebo concluyó que la suplementación con fosfatidilserina produce mejoras significativas en el trastorno por déficit de atención con hiperactividad (TDAH), la memoria auditiva a corto plazo, la memoria de trabajo y el rendimiento mental ante estímulos visuales [73].
La fosfatidilserina utilizada en los primeros trabajos científicos en los que se evaluaba su efecto beneficioso era proveniente del cerebro bovino [74]. Sin embargo, debido a las preocupaciones con la encefalopatía espongiforme bovina, su empleo fue substituido por la fosfatidilserina de fuente vegetal, obtenida de la lecitina de soja. El contenido natural de fosfatidilserina en la lecitina de soja es de cerca del 3%. Para incrementar esta cantidad, se procede a un proceso denominado conversión enzimática que mimetiza procesos enzimáticos naturales. En MINFIRE se utiliza fosfatidilserina de origen vegetal, procedente de la lecitina de soja, con una concentración aumentada del 20%.
El ácido docosahexaenoico (DHA) del omega 3 es el principal ácido graso poliinsaturado del sistema nervioso central y parte constituyente del fosfolípido fosfatidilserina en el cerebro [71]. Por este motivo, un suministro correcto de DHA es esencial para la integridad de las membranas celulares de las células neuronales [75]. Eso se ha evidenciado en un estudio in vivo en el cual se evaluó el efecto benéfico en el organismo de la combinación de fosfatidilserina y DHA. Dicho estudio concluyó que esta combinación puede reducir la concentración de especies reactivas de oxígeno dañinas en el cerebro de manera significativa y puede mejorar las capacidades de aprendizaje y memoria [76].
El DHA es uno de los componentes de otro producto exclusivo de Nutribiolite, el OMEGA 3 + VIT K2 + VIT D3. Este complemento alimenticio tiene 1000 mg de ácido graso puro OMEGA 3 estandarizado al 50% de EPA (500 mg) y al 25% en DHA (250 mg). Recomendamos la suplementación combinada de MINFIRE y OMEGA 3 + VIT K2 + VIT D3. En este link puede consultar un artículo de revisión interesantes sobre las propiedades de los ingredientes del OMEGA 3 + VIT K2 + VIT D3.